“物种”可能不是真实存在的概念?下纲、总目、总科,这无限细分的分类到底是为哪般?
陈光霁 与 孙仲夷 ZJU生命演化研究 2024-04-01 21:41 浙江
(提示:快速浏览完这篇文章大概需要30分钟,带着思考大概需要0分钟。这个问题还有许多未解之谜,如果你有兴趣更深入思考,也许值得投入一辈子。)
引言
达尔文(Charles Darwin, 1809 - 1882) 在《物种起源》中曾言:“迄今为止没有一个定义能够令所有的自然学家都满意;然而每个自然学家在谈论物种时都隐约知道他所指的是什么”。物种的本质一直是生物学和哲学领域的热门议题。生物学家对“物种”的定义长期以来都存在分歧,哲学家则对物种的本体论地位存在争议。然而,作为当今生物分类学中的重要单位,正确理解“物种”概念对生态(考古)调查、环境保护、演化生物学等多个领域至关重要!
虽然在定义上存在着种种挑战,但从亚里士多德对植物和动物的区分开始,延续至现代科学的分类命名法,我们一直在试图对生物体(organism)进行分类(taxonomy)。现代分类系统是基于将生物体组成称为“物种”的单元,这些物种再汇聚形成更大的分类单元,如属、科、目、纲、门等,在生命的嵌套层次结构中连接起来。但是,构成一个物种的要素是什么?为何我们应该将特定的生物群体视为一个独特单元,并为其赋予特定名称?“物种”这一概念究竟是自然客观存在的,还是人为赋予的,这值得更深入地探讨。
Are species real ?
2023年10月,“物种和分类学家的无声灭绝”一文在中文和英文互联网论坛都有引发了广泛的讨论(Löbl, 2023)。分类学家呼吁:“若未能正确鉴定生物并命名,生物学将失去其意义”;旁观者评论说:“分类学的消亡可能涉及科学立场的变迁。作为描述性科学,尽管技术不断发展,但仍然存在其局限性。”;另外一些现实主义者则批评道:“这篇文章只是在‘呼吁’政府提供更多经费给分类学家们而已,似乎并不能带来实质性改变。” 那么,物种分类,是否值得投入更多经费进行研究,物种的概念是否来源于人为主观创造?深入了解这些争议,或许会使你对此有不同的看法。

图1. 生物等级分类阶元示意图(图修改自:https://sciencenotes.org/species-definition-and-examples-in-biology/)
科学术语“物种(species)”一词来源于拉丁文中表示性状或现象的一个词。在试图对生物界进行分类时,人们被迫思考如何合理定义物种的内涵。在讨论争议核心“物种的真实性”时,我们不可避免地会面临两个问题:1. “物种”是否是真实的自然事物(Nature Kind)?2. 如果真实,我们在认知世界的过程中该定义“物种”?真实的自然事物应当如同氢、氧、原子、电子、蛋白质、DNA、生物、基因等一般,具有“服从科学规律”,“作为科学研究的对象”,“反映自然界上真实的事物和过程”的特性,是具有“必要和充分属性的事物”的集合;“物种”这一概念是否具有这样的特性呢?“物种”相对于同其他分类阶元是否有着其独特性呢?
1. 物种是真实存在的生物学实体(Species Are Real Biological Entities)
a. 早期的物种概念
早在亚里士多德(Aristotle)时期(公元前384 - 前322),他就对“物种”这一名词进行了一系列的定义:“物种是真实存在的事物,具有其本质特征。人们能够清楚地将一个物种与另一个物种区分开来,不同物种相互之间不重叠并可进行比较。”。在亚里士多德式的观点中,许多早期的专业术语被运用于对生物体(organisms)进行定义:如属(genus)是一个非常广泛的类别,而物种(species)是指属的内部一类特殊的生物体,可由种差(differentia)所区分。这一套基本概念,被著名的植物学家林奈(Carl von Linné,1707 - 1778)等人沿用并推广,提出了对生物体以拉丁语进行命名的双名***命名法(binomial nomenclature)***,即种名由两部分组成,属名作为第一个词,种加词作为第二个词。种加词一般为描述性词语;如智人(Homo sapines):Homo为属名,译为人(名词),sapines译为明智的(形容词)。

图2. 双名命名法的提出者-卡尔·冯·林奈
这套由林奈主张并推广的双名命名法至今仍然被广泛沿用。然而,在林奈时代,这套命名法仍存在一些问题。例如,由于研究材料通常是死亡和保存不佳的标本,生物学家在对生物进行描述和分类时只能依靠形态学特征。随着越来越多的人将形态学特征作为分类标准,识别和区分新物种不可避免地变得更加主观。
英国著名真菌学家约翰·兰斯巴腾(John Ramsbottom, 1885 - 1974),他在林奈的大量著作基础上整理出了早期的物种概念。早期的物种概念具备三个主要标准:1. 物种是独特的(distinct),独立的(monotypic);2. 物种是不可变的(immutable),天生如此;3. 物种具有纯合繁殖(true breeding)的特性。受限于当时的学术背景,即使在林奈后期的科研生涯中,也出现了许多复杂的理论,试图通过杂交来形成物种;但物种的不可变性(immutable)是18世纪及19世纪初相关研究人员的共识。早期的物种概念受益于大量的博物馆标本,在识别和区分物种时主要侧重于形态学特征,同时也强调了“繁殖”属性的重要性。
物种的不变性与变异性
在强调不变性的物种概念中,物种被视为具有显著特征且不变的实体。物种的归属通过关键性状的确定和相似特征的自动分类来实现。这种观点的根源可以追溯到柏拉图(Plato)时期(约公元前429年 - 前347年)的理念论哲学。柏拉图强调了物种的绝对不变性,认为它由本质(essence)所决定,类似于化学元素的性质。
而在强调变异性的物种概念中,拉马克(Jean-Baptiste Lamarck, 1744 - 1829)认为生物之间存在着不可分割的连续性,而达尔文倾向于认为物种是在极长时间内经历改变和分化的产物。拉马克认为物种只是人为构建的概念,实际上并不存在。然而,有人反驳他过分强调了物种的变异性,而忽视了物种的相对稳定性。相较之下,达尔文认为:“物种只是具有显著特征且相对稳定的变种,每个物种首先是作为变种而存在”(Darwin, 1872)。对达尔文而言,一旦将有机生命视为在漫长时间内经历改变和分化的产物,“物种”这个概念就变得相当任意起来。
b. 生物学物种概念(BSC)
时间追溯到19世纪初,分类学和系统学越来越多的使用博物馆标本之间的形态学差异进行研究,衍生出了自然主义学家和形态学家两大派系。自然主义学家达尔文,提出的自然选择假说极大地推广了“隔离”这一概念,并将“生殖隔离”这一特性阐述得更加具体。
• 隔离是物种形成的必要条件。
• 隔离一般分为地理隔离和生殖隔离两类。
• 长期的地理隔离使两个种群失去交配机会的同时各自积累了变异,导致生殖隔离。
• 生殖隔离是指进行有性生殖的生物彼此之间不能杂交或杂交不育。
• 生殖隔离可分为受精前的生殖隔离和受精后的生殖隔离。

图3. 生物学物种概念的提出者恩斯特·迈尔
著名的演化生物学家与分类学家恩斯特·迈尔(Ernst Walter Mayr, 1904–2005)提出了被大多数现代教科书采用的生物学物种概念(biological species concept,BSC):物种是由相互能够交配的自然群体所组成的类群,这些类群间存在着生殖隔离,也就是说,一个物种是一个能够实现生殖共享的群体(Mayr, 2001)。早在该概念的数世纪之前,就有关于通过生殖隔离来区分不同物种的思想。然而,在许多情况下,确定群体间是否能够相互交配在技术上是困难的,因此分类学家不得不主要依靠形态学上的差异来进行分类。正如达尔文早先观察到的那样:地理隔离是导致生殖隔离的重要因素之一。迈尔在1942年指出,如果某一群体和其亲本物种在地理上隔离,并在隔离期间获得了使其在外界障碍消失后依然能够促进或保证生殖隔离的特征,那么就形成了新的物种(Mayr, 1982)。地理隔离导致的物种形成方式也被称为异域成种,但自然界中仍然存在着大量的同域成种现象。
同时期,同时拥有博物学和遗传学背景的演化生物学家杜布赞斯基(Theodosius Dobzhansky, 1900 - 1975)意识到物种代表的是演化过程中的一个阶段。因此,他并不认为只有地理隔离存在时才会形成新物种,并且认为生殖隔离应该是物种分化的一个副产品(Dobzhansky, 1935)。迈尔及其一些支持者声称生殖隔离是新物种形成的必要条件,否认了同域物种形成的可能性。杜布赞斯基提出了一个颇具前瞻性的观点:“物种并非静态单位,而是演化分歧过程中的一个阶段”(Dobzhansky, 1951)。

图4. 昆虫学家休·帕特森与他的著作《演化与物种的识别概念》(图源:维基百科)
随着生物学物种概念更广泛地被接受,生殖隔离的概念也被更广泛地讨论与总结。在“Evolution and the Recognition Concept of Species”中,它被概括为:包含复杂相互信号和交配及求偶的信号系统,以确保特定配偶能够相互识别的基本功能。昆虫学家休·帕特森(Hugh E. H. Paterson, 1926 - 2019)将其称为特定配偶识别系统(Specific Mate Recognition System, SMRS);物种则被其独特的SMRS定义,新物种的出现(物种形成)即意味着新的SMRS的产生。在广义的生物学物种概念中,物种可以被视为具有共同受精系统的最具包容性的种群。
c. 生物学物种概念(BSC)存在的问题
作为目前最广为接受的物种概念(Species Concept),生物学物种概念(BSC)仍然存在一些问题:
- 博物学家、植物学家和昆虫学家倾向于使用显著的形态特征或纯粹的表型方法来区分物种。特别是在植物中,由于种间杂交现象很常见,生殖隔离并不是一个实用的标准。
- 无性生物和专性孤雌生物缺乏导致配子融合的特定配偶识别系统(SMRS),因此广义的生物学物种概念(BSC)无法严格适用于它们。
- 生物学物种概念只能用于确定同域种群(sympatric populations)之间的生殖隔离,而不能适用于异域种群(allopatric populations)或同域但不同时期的种群。
- 基于群体不断演化的观点,物种群体内并不总是在繁殖上完全隔离,可能存在中间形态(intermediates)和逐渐融合(intergradation),有潜力进行杂交的群体通常不是完全的单系群(Monophyly)。
d. 系统发生学物种概念(PSC)
在1982年,美国古生物学家和鸟类学家乔尔·克拉克拉夫特(Joel Lester Cracraft, 1942 – 至今)提出了系统发生学物种概念(Phylogenetic Species Concept, PSC)。这一概念将物种定义为一个不可简约的群体,其成员来自于一个共同的祖先,并且所有成员都具有一组特定的定义性或派生性特征。克拉克拉夫特提出系统发生学物种概念(PSC)旨在提供一个比生物学物种概念(BSC)更为可行的替代方案。
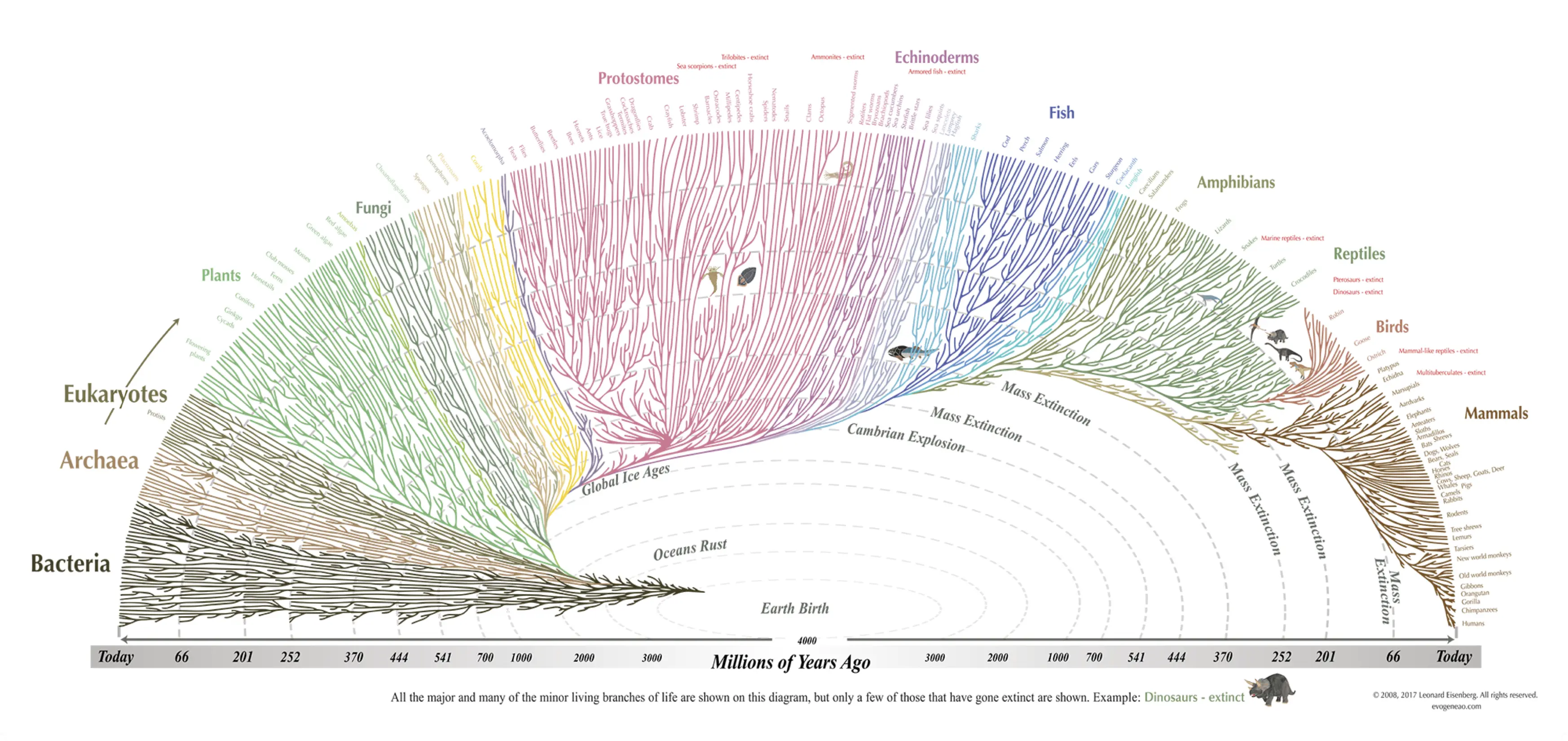
图5. 生命之树 (图源:evogeneao.com)
系统发生学物种概念(PSC)将物种定义为具有共享和独特演化历史的群体。与生物学物种概念(BSC)不同,系统发生学物种概念(PSC)不再将物种之间的繁殖作为唯一的障碍,其限制性更小。即使物种在连续的血统中不断演化,系统发生学物种概念(PSC)也允许对连续的物种进行定义,保留了有性生殖的连续性。在系统发生学领域,已经提出的三个不同的物种概念都以识别系统发生系统中的最小分类单元为“物种”为目标,将物种视为等级降序系统中的最小元素。然而,因为几乎在任何一组生物之间都可以找到微小的差异,在系统发生学物种概念(PSC)下,会倾向于将物种划分为越来越小的群体。
e. 其他物种概念
除了生物学物种概念(BSC)和系统发生学物种概念(PSC)以外,不同研究领域的专家依据自身经验提出了其他的物种概念,例如:
形态学物种概念(Morphological Species Concept, MSC):作为最早的物种概念之一,其定义物种为具有相似形态特征的个体的集合。这个概念被早期博物学家们广泛地应用于生物标本及化石记录等。然而,它不能解决形态差异可能由环境因素引起的问题。
生态学物种概念(Ecological Species Concept, EcSC):这个概念定义物种是在生态位上与其他群体隔离的群体。这个概念强调了物种的适应性和生态角色,但是在实践中很难测量。一个或者一组亲缘关系接近的谱系,占有着与其他任何同域或者从异域演化而来的谱系有着极小不同的适应区间(Turesson 1922)。
***演化物种概念(Evolutionary Species Concept, EvSC)***:Wiley 和 Mayden 基于Simpson的术语,提出了该概念:“由有机体组成的实体,通过时间和空间维持其与其他实体的区别,具有自身独立的演化命运和历史趋势” (Mayden 1997; Wiley and Mayden 2000)。
基因型聚类物种概念(Genotypic Cluster Concept, GCC):定义物种为在基因型空间中形成独立聚类的群体。强调物种的遗传连续性和遗传差异。
……
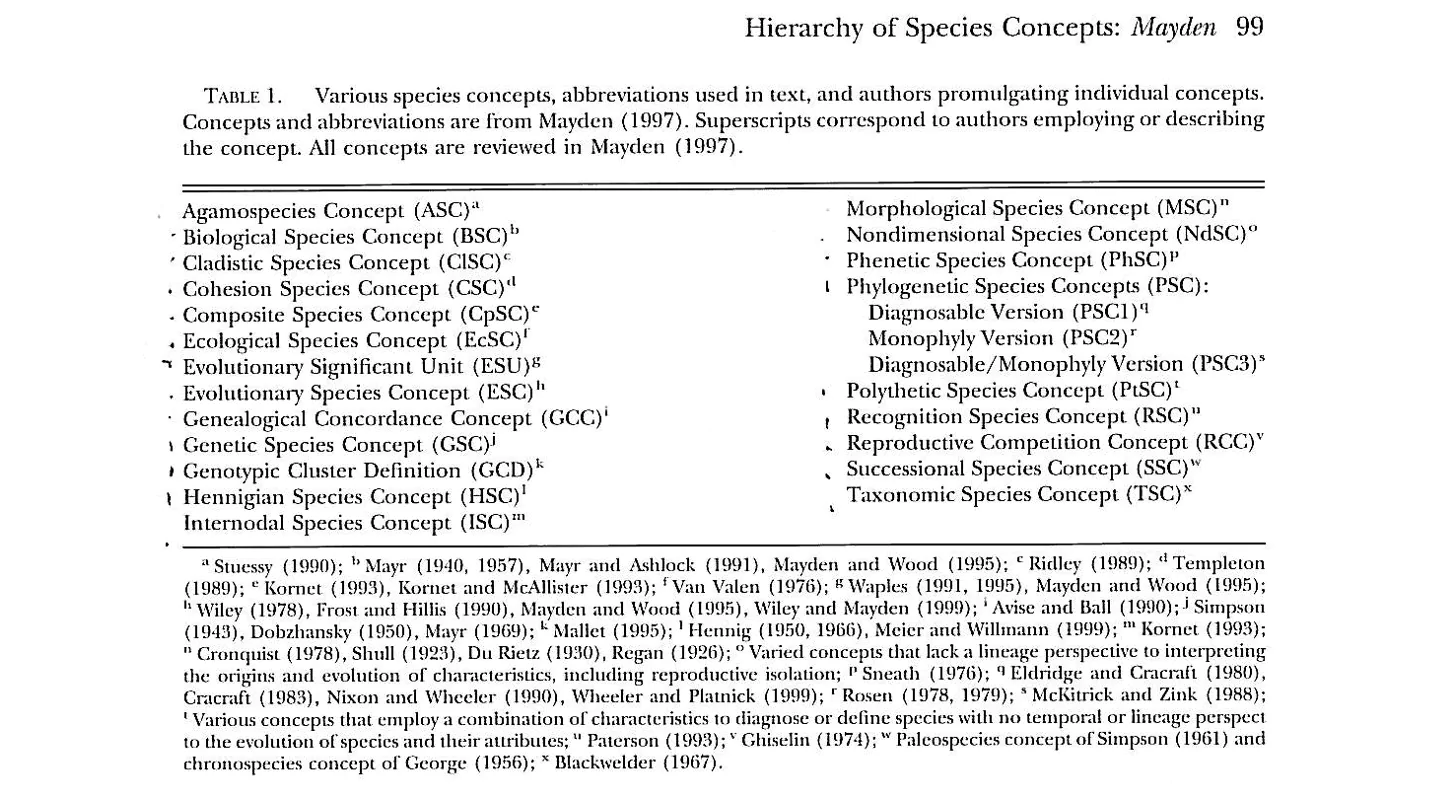
图6. 理查德·梅登(Richard Mayden) 整理的物种概念列表 (Mayden, 1999)
依据鱼类学家理查德·梅登(Richard Mayden)整理的物种概念列表(Mayden, 1999),约有20多种物种概念(species concepts)。依据哲学家理查德·理查兹(RA Richards)在 “The species problem: A philosophical analysis”一书中的区分方法(Richards, 2010),我们可以把这些物种概念大致可以分为三大类:
Process concepts (过程概念类): Biological Species Concept, Recognition Species Concept, Genetic Species Concept, Agamo Species Concept, Ecological Species Concept;这类概念强调“隔离”、“繁殖”这类特性,共同组成了广义范围的生物学物种概念。当然,这类概念也因难以将其应用于自然而受到批评(Richards. 2010)。
Similarity concepts(相似性概念类): Morphological Species Concept, Phenetic Species Concept, Polythetic Species Concept, Genotypic Cluster Concept, Genealogical Concordance Concept;这类概念强调表型-形态上的“相似性”这类特性,这类概念可能是最容易在实际分类认知研究中进行应用的概念,但它会存在“隐存种无法识别”以及“未考虑种内变异”等问题。
Historical concepts(历时性概念类): Evolutionary Species Concept, Successional Species Concept, Paleospecies Concept, Chronospecies Concept, Cladistic Species Concept, Composite Species Concept, Internodal Species Concept, Phylogenetic Species Concept;这类概念是较为新的概念,强调的是生物的“演化历程”及“血缘关系”,并以“物种形成事件”为物种的区分界限。
这些物种概念都试图解决原有的物种概念的一些问题,例如,生物学物种概念不能应用于无性繁殖的生物,不能应用于化石记录,对于是否存在种群之间的广泛杂交,以及对于物种的定义过于严格等问题。然而,每一个物种概念都有其自身的优点和缺点,至今没有一个概念能够完全解决所有的问题。
2. 物种并不真实存在(Species Are Not Real)
在自然世界中,像“电子”、“原子”这样的真实事物具有可区分的自然特性,能够在不同领域内外达成一致意见。在生物学中,存在多种“物种概念”,那么究竟哪一个才是正确认知生物体的合适概念与方法呢?正如理查兹在“The species problem: A philosophical analysis”中提到的:“这种希望形成统一方法的愿望很重要,因为它反映了科学中统一性的价值”。鉴于至今仍无法达成一个共同认可的概念,是否意味着“物种”这一概念并非真实存在,而只是人为创造的抽象概念呢?
a. 生殖隔离?鸟的多次杂交现象

图7. 发生在鸟类的多次杂交现象
2018年“Biology letters”上报道了发生在宾夕法尼亚州的罕见的三种鸟类物种在(属内+属外)多次杂交现象(Toews, 2018)。雌性的森莺属内杂交种(金翅莺Vermivora chrysoptera和蓝翅莺Vermivora cyanoptera 的属内杂交种)与雄性的橙尾鸲莺属的栗胁林莺Setophaga pensylvanica成功交配并繁殖,产生了后代:一只雄性的跨属杂交种。这只雄性跨属杂交种唱的歌曲像橙尾鸲莺属的栗胁林莺S. pensylvanica,但形态特征酷似森莺属Vermivora。这是一种罕见的多次跨种又跨属杂交现象,挑战了物种之间的“生殖隔离”的这一标准。这似乎表明我们对“生殖隔离”、“物种形成”和“物种概念”的理解可能需要进一步的深化和修正。
b. 生殖隔离?环物种现象
另一个有趣的自然现象,也让“物种”这一概念变得不那么真实:“环物种”(Ring species)。环物种是一类非常有趣的生物学现象,环物种是指一个物种因地理区隔(如湖泊、山岳、峡谷)等因素而沿着该区隔繁衍产生多个亚种,各相邻亚种之间有着连续性的基因变化。然而,当此一连续亚种之首尾物种亦相邻时,首尾两亚种却因差异太大而不进行杂交繁殖。这就形成了一个“环”,在这个“环”中,每个亚种都可以和相邻的亚种进行杂交,但是环的两端的亚种却无法进行杂交。
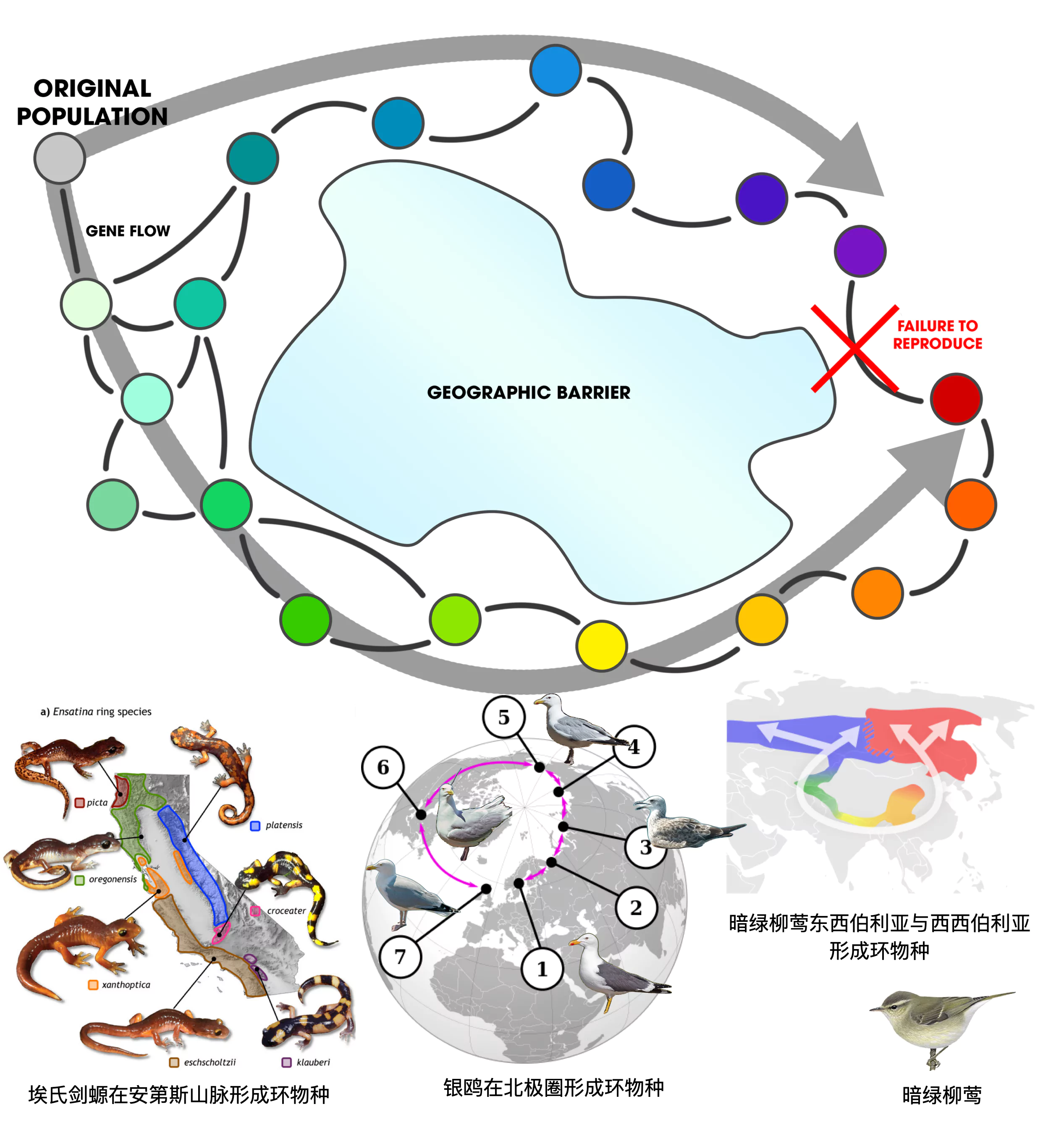
图8. 环物种形成模型(埃氏剑螈、银鸥及暗绿柳莺的环物种现象) (图源:维基百科)
环物种现象对于物种之间的“生殖隔离”,“无法杂交”的标准提出了质疑。传统上,我们认为如果两个群体不能进行成功的杂交,那么它们就应该被认为是不同的物种。然而,环物种的存在表明,即使是在同一物种内的不同亚种之间,也可能存在无法进行杂交的情况。这就使得“生殖隔离”这个标准变得模糊。
环物种现象,一方面被用于提示“物种形成”的一个模型,即生物的演化是一个复杂的过程,物种的形成和生殖隔离可能比我们想象的要复杂得多;另一方面也意味着基于“生殖隔离”的“物种概念”的标准需要进行革新。
c. 人为创造的“物种”概念?
前文提到的各种“物种概念”的提出者,都是基于物种“Species”是一个真实存在的类群,不同的物种概念都极力地找寻将生物体区分成不同“物种”的关键相似性和关键差异点,但十分可惜的是,迄今为止都没有一个统一且令人完全信服的物种概念。如果我们的目标是理解地球上的生命,大多数的概念在应用于细菌等无性生物体(毕竟它们真的是地球上的主要存在者)时会出现各种棘手的问题。当然,在植物、真菌和部分动物中也存在无性繁殖,这些概念也会束手无策。
从“形态”到“生殖隔离”,再到“共同祖先”,物种概念的不断革新,仍无法令各个学科达成统一的共识,一部分人便主张“物种实际上并不真实存在”,并将物种概念问题归结为关于一种任意的分类系统的辩论。当然,物种问题的持续争论,也表明许多人并不愿意如此轻率地一笔带过。
另一方面,从真实的自然事物出发(Nature Kind),“物种起源和灭绝”似乎暗示着“物种”在某种程度上是真实存在的。因为抽象的类别(比如“氧原子”)是不会灭绝的:如果所有的氧原子不知何故消失了,氧气不会“灭绝”,因为只要形成了一个新的具有八个质子的原子,氧气就会再次出现。而物种却无法如此:当最后一只渡渡鸟死亡后,该物种就灭绝了。即使是从恢复的渡渡鸟细胞克隆出的生物,也不会是渡渡鸟,因为它将在与从前极度不同的环境中发育和存在,成为不同于过去生物的新的生命体。
3. 物种不是独特的生物学实体(Species Are Not Uniquely Real Biological Entities)
在讨论“物种不是独特的生物学实体”这一观点之前,需要介绍“PhyloCode”这一新兴事物,它与这一观点密切相关。传统分类法中的命名和分类根据形态特征和共同祖先的共有特征(存在一些问题) : 1. 命名和分类不一定能很好地反映物种的真实演化关系。2. 由于物种形态特征存在连续变化的特点,分类单元之间的边界通常模糊不清,导致分类的主观性和不一致性。PhyloCode的诞生便是为了解决传统分类法的上述问题:
a. PhyloCode (国际系统发生学命名法规)
国际系统发生学命名法规,简称PhyloCode,是一套新兴的基于系统发生,用于管理系统发生命名的正式规则。PhyloCode的当前版本专门设计用于规定类群(clades)的命名。PhyloCode与国际系统发生学命名法学会(ISPN)相关联,并于2020年提供了《Phylonyms》命名手册,根据PhyloCode法规确定了300个分类名,为不熟悉该法规的人提供了示例。RegNum是一个用于注册类群名称的相关在线数据库(截止2023年12月已注册603个类群)。

图9. 《PhyloCode》与《Phylonyms》分别于2010年、2020年发表 (图源:网络)
PhyloCode 的理论基础是在1990年代由 凯文·德·奎罗斯(Kevin de Queiroz) 和 雅克·戈提耶(Jacques Gauthier) 在一系列论文中基于“关于分类群名称可以根据系统发生树的一部分来定义”的建议之上进一步提出的。PhyloCode通过提供决定哪些名称和定义的关联被视为已建立的规则来规定系统发生命名。PhyloCode仅管理类群的命名,不管理并系群或多系群,且只允许使用标本、物种和突变作为指定物(锚)。
与基于阶元(如属、科等)的命名法规不同,PhyloCode不需要使用等级(rankless)。基于等级的法规使用等级和在许多情况下的类型标本或类型亚类群来定义类群。在系统发生命名法下,类群(Clade)的划分是基于系统发生(即,祖先和后代)的定义和使用指定物(如物种、标本、突变)来表示其组成的实际生物。
PhyloCode使用严格的系统发生学概念,例如节点(node)和演化支(clade)来定义分类单元。这样,PhyloCode能够提供一个更直观、更准确的分类系统,能够更好地反映物种之间的亲缘关系和演化历史。举几个例子:
- 鹦鹉雀类鸟:Psittacopasseres,在RegNum里注册号为718,其官方定义:“包含了灰鹦鹉Psittacus erithacus Linnaeus, 1758 (鹦鹉目鸟类) 和 家麻雀Fringilla domestica (现称为 Passer domesticus) Linnaeus, 1758 (雀形目鸟类) 的最小冠群。”。 [Verbatim Definition The least inclusive crown clade containing Psittacus erithacus Linnaeus, 1758 (Psittaciformes) and Fringilla domestica (now Passer domesticus) Linnaeus, 1758 (Passeriformes).]
- 哺乳动物:Mammalia,在RegNum里注册号为220,其官方定义:“包含了智人 Homo sapiens Linnaeus 1758 (胎盘动物)、有袋负鼠 Didelphis marsupialis Linnaeus 1758 (有袋目) 和 澳洲针鼹Tachyglossus aculeatus Shaw 1792 (单孔目) 的最小冠群”。 [Verbatim Definition: The smallest crown clade containing Homo sapiens Linnaeus 1758 (Placentalia), Didelphis marsupialis Linnaeus 1758 (Marsupialia), and Tachyglossus aculeatus (Shaw 1792) (Monotremata).]
- 哺乳形态类:Mammaliamorpha,在RegNum里注册号为222,其官方定义为:“包含 智人 Homo sapiens Linnaeus 1758 (哺乳纲)和 三瘤齿兽 Tritylodon longaevus Owen 1884(三齿兽科)的最小支系群”。[Verbatim Definition: The smallest clade containing Homo sapiens Linnaeus 1758 (Mammalia) and Tritylodon longaevus Owen 1884 (Tritylodontidae).]
b. 为什么物种不是独特的生物学实体?
正确定义的物种是真实存在的实体,但不是唯一的真实存在;所谓的“物种问题”实际上是分类问题的一个特殊案例。
Species “properly defined are real entities, but not uniquely real” and argues that the “so-called ‘species problem’ is really just a special case of the taxon problem”. –Brent Mishler
UC Berkeley的植物学家Brent Mishler依据PhyloCode的无等级(rankless)的思想,进一步讲其激进地阐述其观点:“物种Species”这个分类阶元可能并不具有其独特性,我们不必拘泥于等级划分思想为物种进行命名。Brent Mishler 认为“phyloCode 基于生物的演化历史来进行命名和分类,不再需要等级制度,这种方式更能体现生物的实际演化过程,更加准确合理”。
这些内容或许有些激进得让人难以接受,如果我们可以跳出原有的教科书思维,真实地从双方的角度去思考“物种问题”。Mishler教授所倡导的无等级(rankless)思想,很大程度地消解了,在传统的分类法则下因为过于依赖人为制定的等级制度而造成的混乱以及其不准确的结果。该思想并未触及“物种”这一划分概念的真实性,但动摇了其独立于“属”、“科”、“目”这类等级的独特性(过去定义这些独特性是通过形态,生殖隔离的繁殖特性,或者共同祖先来进行区分的)。Brent Mishler接受物种之间的分化是真实存在的,但他认为“在何时将两个分化后的分支划分为不同的物种目前是没有固定不变的标准(与“属”与“属”之间,“科”与“科”之间一样并没有固定的划分标准)。
达尔文明确提出物种的概念是一个相对随意的划分点。在《物种起源》中,达尔文反复提及物种等级并不是一种客观必然存在的区分标准,而是基于经验和专业分类学家的判断来确定的。虽然物种概念不断革新,但其“人为区分”的属性只是减弱到了最低,并未完全消失。即使是系统发生学物种概念(PSC)观点中强调的“最小可区分”的特性,也依然需要特定系统学家的主观判断来到底多大程度地不同才能作为区分和识别的标准(这些判断依然是相当主观的)。Cardiff University的昆虫学家Michael F. Claridge认为在确定物种分类群的界限方面,系统发生学物种概念(PSC)和广义生物物种概念(BSC)之间的实际差异不是很大。
以无等级的思想,运用PhyloCode的命名规则,或许在早期会引起大量的混乱和矛盾;但随着更准确的系统发生关系的明晰,将会获得一套更直观、更准确的分类系统,能够更好地反映物种之间的亲缘关系和演化历史;在教学、研究、与其他专业领域的科学家交流以及与社会讨论中会更加方便与准确。
4. 讨论
1. 物种的真实性?Are Species real?
回顾历史,从17世纪开始,随着科学技术的发展和海运的兴起,博物学家们从世界各地搜集到了大量的动物、植物和化石标本。由于没有一个统一的命名法则,各国学者都按自己的一套工作方法给当时的植物命名。从早先公元1600年已经认知的约6,000种植物,到后来公元1700年前后又新发现了约12,000个新物种。那时,各国学者用冗长的“短语”命名动植物,出现了相同的动植物却有不同的名字、而一些不同的动植物却又同名的怪现象。这种现象影响了人们对动植物的统一认识,影响了国际学术界的交流。于是,迫切需要对这些生物物种进行科学的分类。存在着两大类:各分类类群的物种标准不一致,以及各个物种概念的标准不一致的问题。
长期以来,物种的分类是基于性状相似性的比较,是对形态的、生理的、遗传的、行为的性状数据进行评估,利用证据进行推导的过程。虽然常常不能直接观察到物种的生殖隔离或物种分化,但可依据观察到的各种证据,如形态差异,对物种地位进行推论。有条件时,利用生物学的手段对这种推论进行验证,最终确定物种的形态学标准。分类单元(或个体)不是因为其相似而构成同一单元,而是因为其属于同一单元而相似。生物分类学家将形态差异的类型和程度视为生殖隔离的外在指标,用作进行推论的证据。
关于“物种问题(species problem)”的讨论,种群特征、连续性特征和可演化特征是造成这一系列争论的源头。在生物学研究中,物种是一个颇有争议的概念。从达尔文时代开始,物种的概念一直在演化。从形态种,到生物种,演化种概念的演化,是从模式种概念向种群概念的转化,即从静态的种的概念向动态的种的概念转变。于笔者而言,“物种”这个分类阶元与“物种”以上分类阶元存在其独特的意义:物种是一个可以随时间而变化的个体集合;物种是生物多样性中三大多样性:遗传多样性、物种多样性和生态系统多样性的重要基础。
我们在“物种”这个问题上,追求概念/原理的普适通用性反映的是科学中统一性的价值。通用性的体现从最早先的基于“形态”差异、到后来的基于“生殖隔离(繁殖特性)”,再到“共同祖先”这一特性。这些都是早期博物学家、植物学家、昆虫学家、动物学家、生物学家等认知“物种”,定义“物种”,追求真理的关键点。如果“物种”这一概念是人为创造,“物种并不真实”,这似乎也不会改变分类学家利用各种主观的关键点对于生物体进行等级划分与命名。但倘若“物种”这一概念真实存在,即使目前这一关键点(用于区分“物种”)目前并未达成共识,通过一代又一代的观察与总结,这个关键点必然会被找到。
2. 种以上分类阶元是否有生物性基础?by 孙仲夷 & 陈光霁
在传统的分类法中,分类阶元从物种(Species)、属(Genus)、科(Family),到更大的阶元如目(Order)、纲(Class)、门(Phylum);在我们探讨物种(Species)概念的真实性之后,考虑到物种内部相对缺乏生殖隔离的生物特性,其他的分类阶元可能并不直接具备真实性这一特征,因为它们缺乏遗传物质的自由交换。然而,“种以上的分类阶元是否具有生物学基础?”这一问题也值得我们进行探讨。传统的分类体系为理解生物体的多样性提供了一个很好的组织框架,帮助我们更好地理解和研究生物体之间的关系。在向比“物种(Species) ”更高的分类级别迈进时,如属、科、目等,这些阶元的界定通常涉及更多的形态学、生态学和演化关系。
基于生物的遗传与演化而言,“物种(Species) ”是一个有特殊地位的分类单元。由于物种内部相对无生殖隔离,祖先种群内的广泛杂交使得种内不同个体的系统发生关系总体上呈现出复杂的网状结构;对于不属于同物种的生物体而言,生殖隔离会使它们的系统发生关系基本呈现出复杂的树形结构(在一定程度上也可能会受到跨种杂交、基因渐渗等现象的干扰而呈现零星的网状结构)。因此,对于物种以上的其他分类阶元,我们需要在生命之树(Tree of life)所假定的树形结构的基础来考察它们的划分是否具有生物学基础。
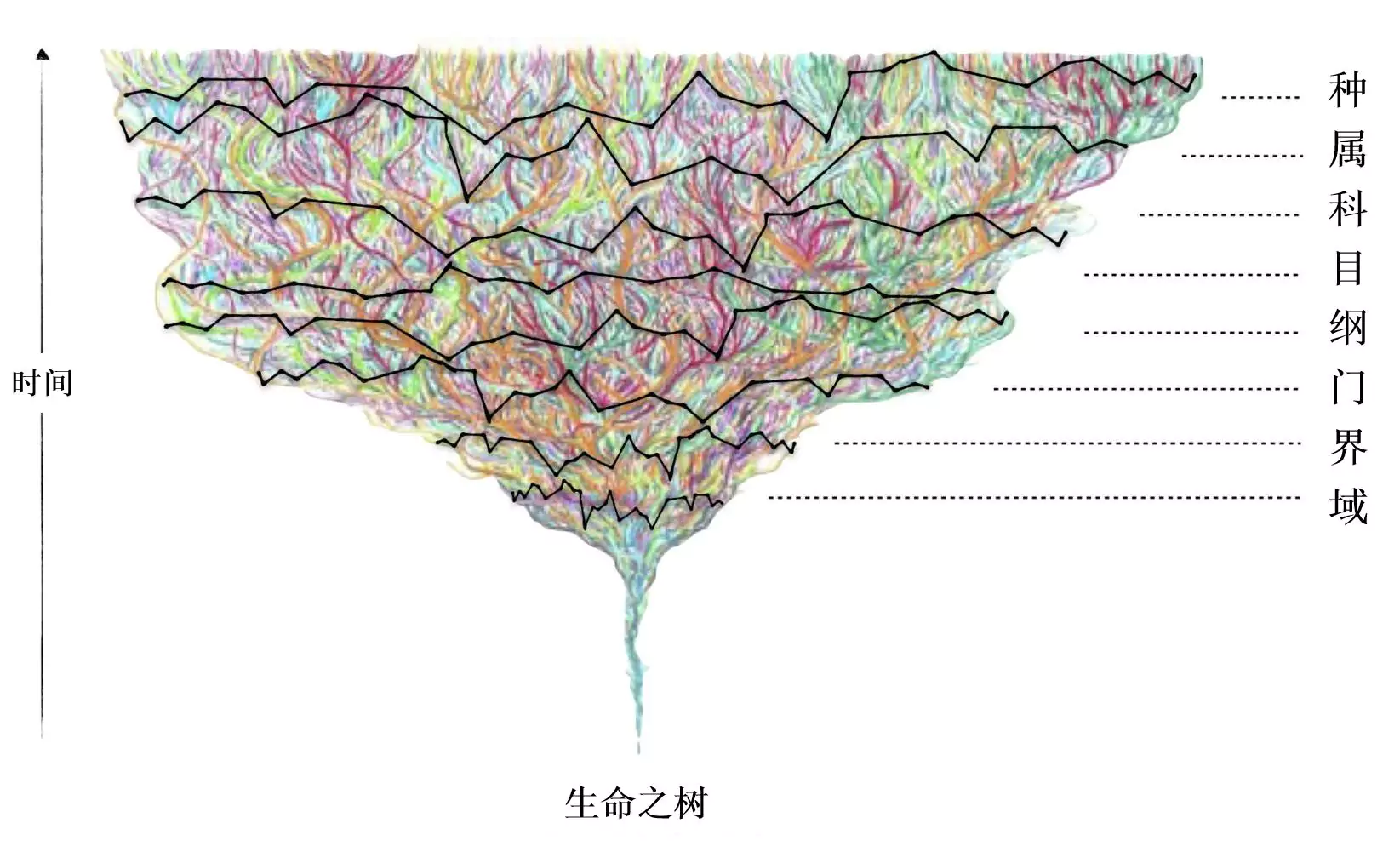
图10. 生命之树上的生命体(彩色线条)与传统分类法的分类阶元(黑色线条)的示意图。
(图修改自:Karen Klitz from Swartz and Mishler, 2023.)
2.1 划分方式的单系性
随着分子生物学技术的发展,特别是基因组学和系统发生学的进步,生命之树(Tree of life)的构建出现了可能。但由于生物体之间的演化关系可能呈现出连续性,而不是清晰的分隔。在某些情况下,物种间的界限可能模糊不清,可能存在相互重叠或渐进过渡的情况,这就使得对不同分类阶元的界定变得更加复杂和模糊。***在系统发生树结构下,我们希望每一级的分类阶元都应当代表一个完整的独立分支,即单系群(*Monophyly)**。大部分的纲(Class)是单系群。例如,我们所熟知的哺乳动物纲(Mammalia)就是一个单系群,它包括了所有哺乳动物的共同祖先及其所有后代。
但事实上,基于各种形态学、生态学和地理分布等描述性表型特征的传统分类方式不能完全满足形成单系群这一要求。例如,根据过去的传统分类方案,爬行纲被定义为“既非鸟类也非哺乳类的羊膜动物”,因此爬行纲(Reptilia)包含了蜥蜴、蛇、龟鳖和鳄鱼等。然而,爬行纲物种的共同祖先的所有后代还包括所有鸟类和非鸟恐龙,因此传统爬行纲并非单系群;一些分类学者提出了重新定义爬行纲的想法,他们希望将鸟类以及非鸟恐龙包含进来,以形成单系群,或者使用包含非鸟恐龙和鸟类的蜥形纲(Sauropsida)来替代过去爬行纲(Reptilia)的定义。从系统发生关系的角度来看,传统的七阶分类方式并不是很理想,有些类别的命名因其数百年的历史惯性而显得积重难返。
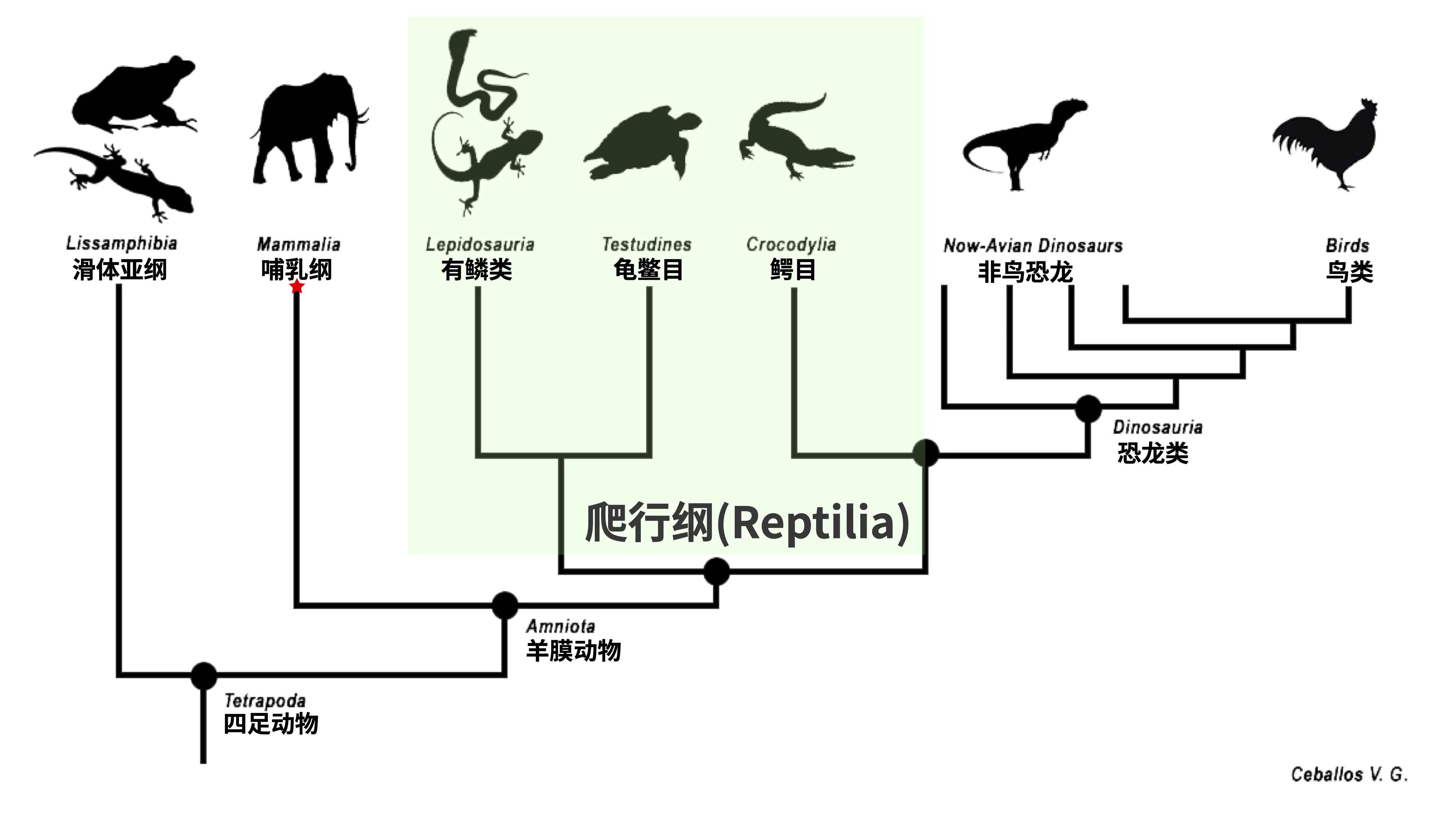
图11. 传统分类方案中的爬行纲并非单系群(图源:Ceballos V. G.)。
然而,为了有效理清物种多样性的规律和起源,我们又不得不设立各种类别概念对某个或大或小的物种集合加以描述。这时候我们需要在传统的门类单元之间插入更多灵活的划分方式,例如,人们将鸵鸟、鸸鹋等所在的四个目的鸟类定义为平胸总目(Ratitae),因为它们都有扁平的胸骨,无龙骨突,翅膀退化,丧失飞行能力等显著特点。但在最新发布的鸟类系统发生关系中,平胸总目并非一个单系群,分布于拉丁美洲的会飞的䳍形目(Tinamiformes)鸟类与平胸总目鸟类中美洲鸵鸟目(Rheiformes)应当是更为近源物种发生关系。因此,包含了,并非单系群的平胸总目与䳍形目(Tinamiformes)鸟类的古颚下纲(Palaeognathae, 也称古颚总目)应运而生。

图12. 古颚下纲鸟类系统发生关系。(Josefin Stiller 和 陈光霁绘)
类似的还有超门(Superphylum)、亚门(subphylum)、小纲(Parvclass)、下目(infra-order)等无数具体概念,但这种细致的类别划分同样要求其内部的所有物种共同构成一个单系群。也就是说,该类别的生物所共享的特征必须继承自它们的共同祖先,而非趋同演化所导致的,这样的特征被定义为同源特征。因此,我们不能因为海豚和蝙蝠都有回声定位的功能而单独地将它们化为一个类别,因为它们的这种生物特征是各自独立演化的结果。
因此,基于演化的分类系统必须反映其产生的自然秩序。也就是说,各种各样的生物特征,从基因序列到复杂表型,它们都要受到系统发生关系的支配和约束,自然选择虽然能改变方方面面的生物性状,但不能给现有的地球生物带来一套全新的遗传密码,或者给现生动物替换出一套截然不同的神经系统。这使得我们充分相信,那些高度保守的基础特征是所持有者继承自它们的共同祖先的结果。如果像脊索、脊椎这样奠定一个门类的主要特征可以独立地多次起源的话,那我们由此定义出的脊索动物门、脊椎动物亚门就毫无演化意义了。
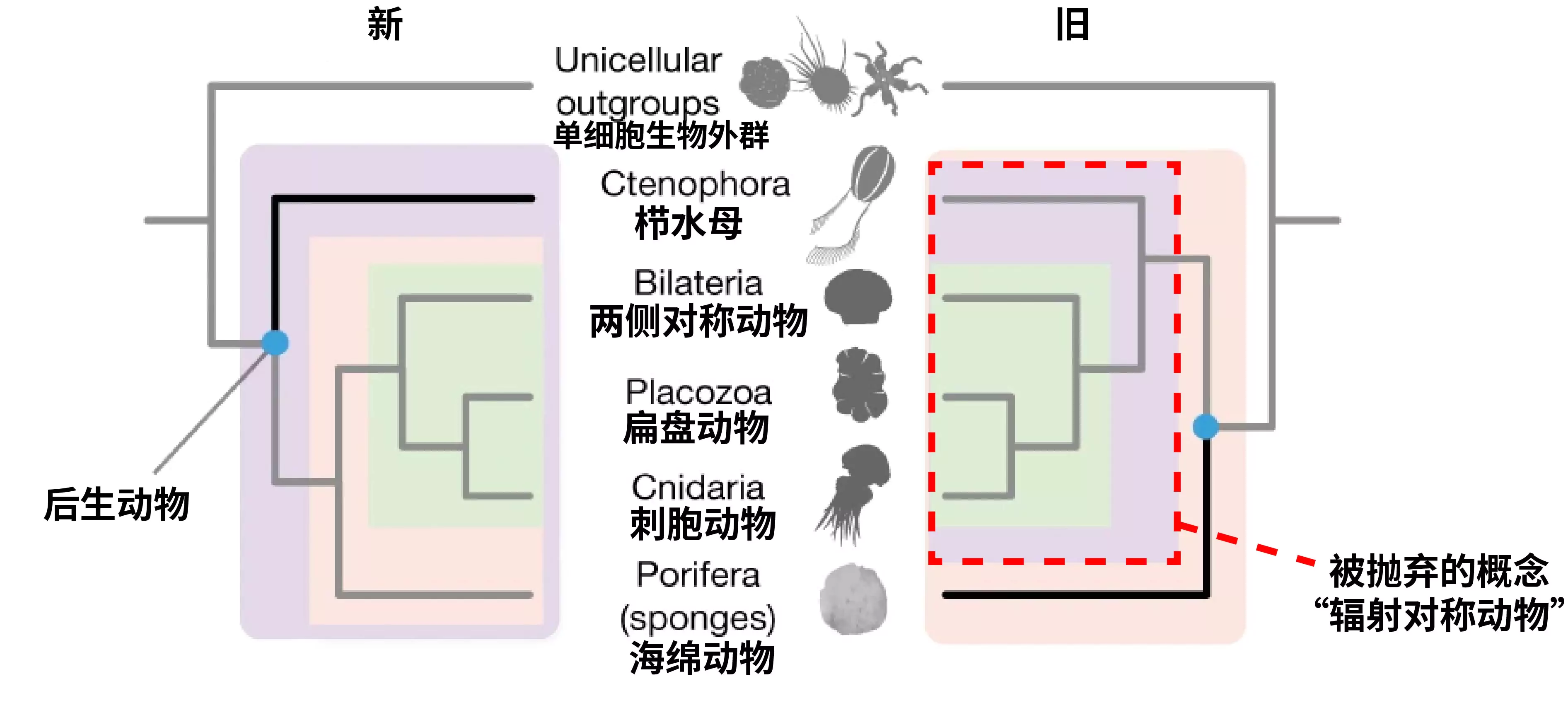
图13. 后生动物主要类群的系统发生关系示意图。左图为新的系统发生关系,支持栉水母为其他后生动物的姊妹裙;右图为以往的系统发生关系,依据辐射对称提出的辐射对称动物(已逐渐被抛弃的概念)。(编辑自https://www.nature.com/articles/s41586-023-05936-6)
与此同时,我们也会不断修正一些认知,例如过去我们根据辐射对称的特征将水母等刺胞动物与栉水母生物划分为辐射对称动物,而基于基因共线性的证据显示栉水母应当是其他所有后生动物的姊妹群,因此无法与刺胞动物划为同一门类,于是近年来,“辐射对称动物”这一概念逐渐被学术界抛弃。从整体上来看,单系性是每个分类阶元都追求的基本原则,但接下来我们会看到,不同的具体分类阶元反映出不同层次的生物共性或特性。
2.2 林奈双名法以及属的意义
当你第一次吃到黄瓤西瓜的时候,你一定会好奇,为什么世上还能有这样的西瓜?有务农经验的长辈这时候也许会解释到“它跟你经常吃到的西瓜不是同一个品种”。换一种更科学的表述方式,我们可以说这种西瓜跟大家经常吃到的红瓤西瓜有着系统性的遗传差异。对于这种不常见的西瓜,我们用“黄瓤西瓜”或“西瓜·黄瓤”来表述就能很容易指出它的与众不同之处来加以区分。
类似的情况在演化研究中再常见不过,例如非洲和亚洲都有体型巨大的大象,但二者又在体型、耳朵大小、象牙长度、趾头数量等方面存在系统性差异。因此,伟大的古生物学家乔治·居维叶(Georges Cuvier,1769-1832年)首次提出将现存的大象分为非洲象和亚洲象两个物种,后来的研究又进一步将非洲象分为非洲草原象和非洲森林象。我们从这里可以看出,演化与生态学家在研究物种多样性和生态功能的时候,绝大多数情况下并不关心两个地理隔离的种群是否形成严格意义上的生殖隔离,而是只要发现了一系列有遗传与生态层面研究意义的,稳定的种群间表型差异,这时候就有必要从相对广泛的生物模式中定义出更具体的物种(Species)。也就是说,在基本认知上,我们可以很容易区分大象这一类生物,但系统性研究要求我们区分出更丰富的遗传演化差异,于是就诞生了林奈双名法。
在林奈之前,物种划分是十分混乱多样的,每个植物学家或动物学家可能都有一套自己的划分方式和命名标准。尤其是为了能全面细致地描述一个物种的特征,当时很多人采用多段词汇堆叠的方式来给物种命名,即多名法(polynomial nomenclature)。随着人类定义出的物种越来越多,这一度带来巨大的混乱。此时林奈引入了一个关键的新思想,即名称的作用只是给一个物种一个独特的标签罢了,大家能知道具体指代的物种是啥就行。于是林奈用两个单词的组合来命名每一个物种,第一个单词代表属名,第二个单词代表种名。只要能保证唯一性,哪怕种和属名都不是精准的特征描述也没问题。例如,鹦鹉科的一只鸟被命名为Psittacula alexandri,意思是“亚历山大的鹦鹉”,以亚历山大大帝的名字命名,亚历山大大帝的军队将东部长尾小鹦鹉引入希腊。
林奈双名法比过去的多名法更容易记忆和使用,并最终取代了它们。这种命名方式历经数百年,直到今天仍能稳定使用下去,这表现了其广泛的科学适用性。
在这套体系里,属(Genus)的作用是至关重要的,因为只要框定了具体属的范围,在此基础上对物种加以区分反倒是一件相对清晰明了的事情。也正因我们无论研究动物植物微生物的时候都普遍使用双名法,属的划分方式必须遵循以下几条十分关键的基本原则:(1)单系性,即一个祖先分类阶元的所有后代被归在一起(即系统发生分析应清楚地证明单系性和作为单独谱系的有效性);(2)合理的紧致性,即属不应该被不必要地扩展;(3)独特性,即符合演化相关的标准或大致共识,即需要从生态学、形态学或生物地理学多方面综合评判。
我们以人类与黑猩猩之间的关系为例,来阐释这一问题。尽管在现今的动物界中,人类和黑猩猩保持着完全相邻的姊妹群体关系,但它们分离的时间已经长达约五百万年。这段时间足以使它们之间形成了稳定的生殖隔离关系,几乎没有任何基因交流。因此,数百万年以来,它们在形态、行为和习性等诸多具体特征上逐渐产生了显著可区分的差异。将它们强行合并到一个属内,将给演化和生态研究带来许多不必要的困扰。而着眼于人属内部,虽然现生的“人类”只有我们智人(Homo sapiens)一个物种,但是尼安德特人(Homo neanderthalensis)等历史上存在过的其他人属物种都曾与智人祖先产生过复杂的基因交流并深刻影响了现代智人的遗传特征,它们的生殖隔离并不严格。而大量的基因组测序分析则向人们揭示了属内物种杂交事件的广泛存在,甚至直接决定了很多新物种的形成。
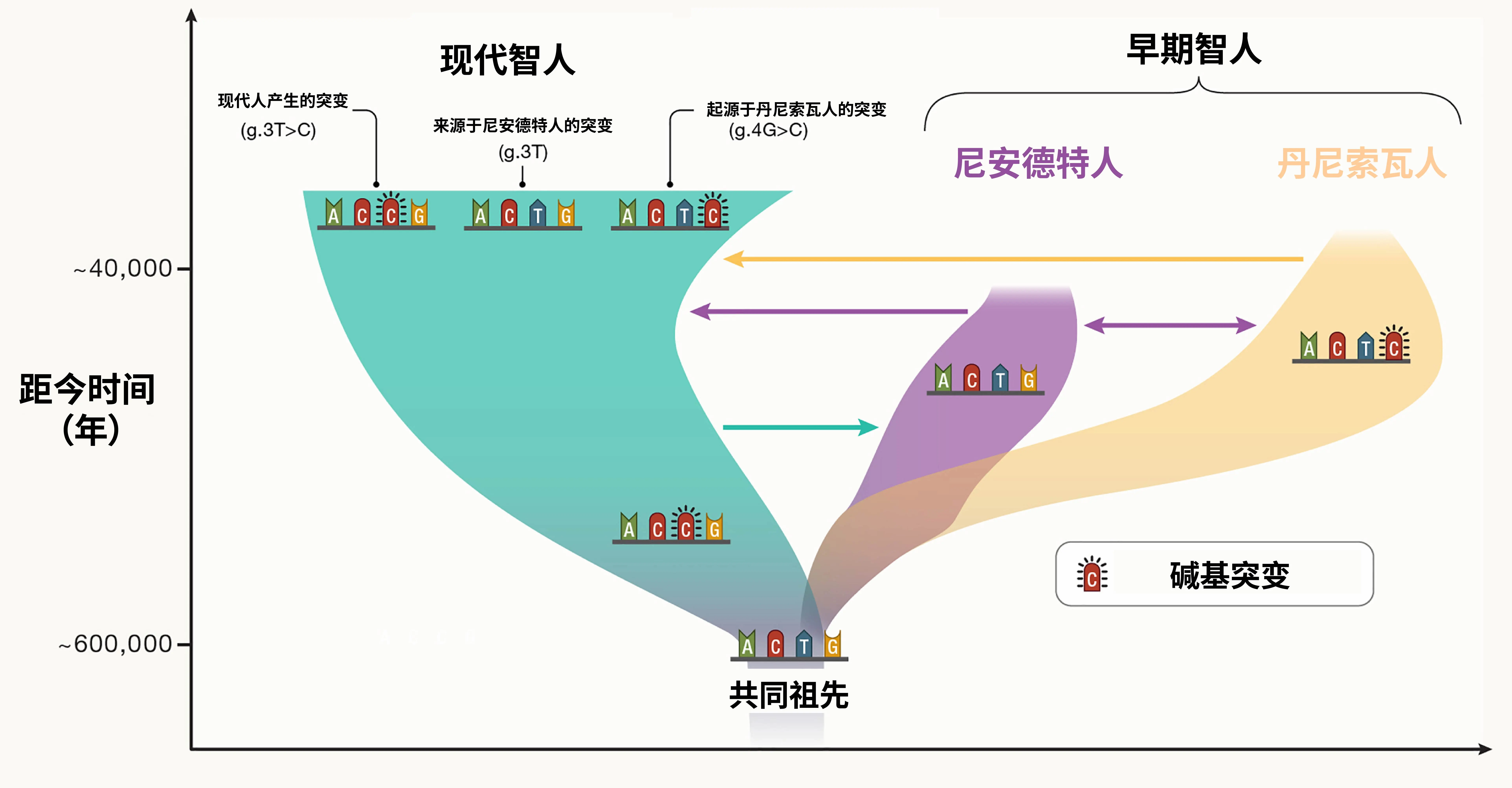
图14. 现代智人与早期智人的基因交流(图源:https://doi.org/10.1016/j.cell.2023.12.029)
总之,“属”这一概念虽然跟“种”一样做不到尽善尽美的定义,对演化生物学研究来说是十分关键的,它往往反映的是人们普遍认知上对于物种的基本划分方式,就像我们谈到人、老虎、狮子、大象、犀牛、猕猴等常见概念的时候,其实都在生物分类学上完全对应着一个具体的属。换句话说,普遍意义上的生殖隔离大多介于属和种之间,一个属的物种类群要保持足够长的独立演化时间才能被人们接受。
我们可以看到,“属”是一个比较严肃的概念,一个新的生物属一旦被鉴定出来往往会在此后数百年维持不变(属的名字可能会发生变更),但种的划分却相对更加灵活多变,例如将过去定义的一个物种内两个亚种重新区分为两个独立物种是很常见的事情。从遗传独立性的角度讲,属是一个比物种更重要的概念,它在系统发生树中更能代表那些几乎互不交叉的基本末端节点。也就是说,自属往上的系统发生关系,整体上能基本保障树状结构,而从属到种的系统发生关系则往往表现出有内部交叉的网状结构。
2.3 纲、目、科的生物学基础
从属往上的分类阶元依次是科(Family)、目(Order),以及纲(Class)。分类学家在描述或识别一个科、目或纲的阶元时没有硬性的规则需要遵循。也就是说,这些等级的划分方式在历史上经常发生频繁的变动。例如“纲”的概念在过去曾承担着描述最广泛的共性分类特征的功能,但该功能被德国生物学家恩斯特·海克尔(Ernst Haeckel,1834年-1919年)提出的“门(Phylum)”的概念所取代。
因此我们今天看到的科、目、纲主要是基于分类学家的历史性努力而逐渐固定下来的,所以对于不同生物门类而言,一个“科”所涵盖的遗传多样性和演化时间往往截然不同且毫无可比性。例如,隐翅虫科(Staphylindae)有上千个属,超过63000个现存的已知物种;而人科(Hominidae)只有4个属,8个现生物种。而且人们在纲、目、科之间往往插入诸如下纲、小纲、总目、下目、亚目等数十个相对不常见的阶元。而这些不常见的阶元之间也经常存在变更的情况,如前文提到的平胸总目到古颚下纲的变化过程。
虽然具体的划分方式五花八门,但逐步分类的过程带来了人们对于生命演化逻辑的理解。最基础的就是前文提到的继承自单系共同祖先的同源性特征。而不同的同源特征的产生顺序就反映了演化的基本逻辑。例如,老鹰所属的鹰形目(Accipitriformes)与猫头鹰所属的鸮形目(Strigiformes),在过去的认知中鹰形目是所有陆鸟类的基部类群,而鸮形目则是晚近出现的独立类群,因而老鹰与猫头鹰所具有的肉食性被认为是陆鸟类祖先特征,但依据最新鸟类生命之树,老鹰与猫头鹰应当属于近源物种,共同所属鹰形总目(Accipitrimorphae)鸟类。这样的重新划分使得对于老鹰和猫头鹰的肉食性这一食性的起源的认知可以重新被改写为仅在鹰形总目鸟类的最近共同祖先里才出现的,陆鸟里其他肉食鸟类可能是独立出现了食肉习性。
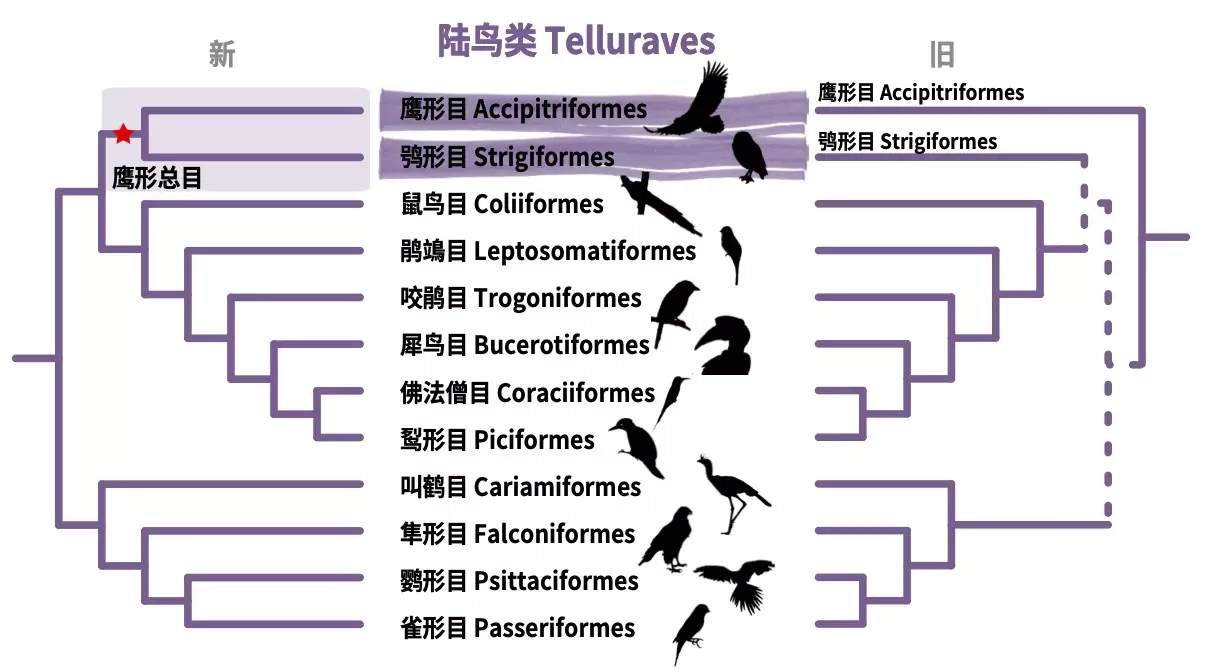
图15. 鹰形目与鸮形目亲缘关系的重新划分示意图。左图为新的系统发生关系,支持鹰形目与鸮形目共同组成鹰形总目,右图为以往的系统发生关系,虚线表示新旧系统发生关系的之间的变化。(Josefin Stiller 和 陈光霁绘)
同源特征也可能因退化而消失,例如蛇类等少数爬行动物分支不具备鸟类与爬行动物普遍拥有的四肢结构,但来自古生物化石和胚胎发育的证据都能证明它们是在后续演化历程中丢失了祖先曾有过的四肢发育能力。但我们必须要找到丢失同源性的明确证据才能把不具备更大类群共性特征的某个小类群(蛇下目)放到大类群(爬行纲)的内部。
总的来说,我们在纲、目、科及其内部插入大量其他分类阶元的目的是理解系统发生的真实过程,而是否意味着存在某些生理或者性状特征使得这一类群物种具有适应性优势或巨大的演化潜力(evolvability),或者足以使得其区别于其他类群,这仍有待商榷。例如上述的蛇下目,均具有相同的四肢退化特征。而在传统偶蹄目(Artiodactyla)中,也因新的证据证明了不具备偶蹄特性的鲸豚类与偶蹄目中河马为近源物种,“偶蹄”这一生理特性所代表的偶蹄动物被认为是无效的并系群分类,因此引发了新的分类定义:鲸偶蹄目(Cetartiodactyla)。在这些中间尺度的分类阶元内,各种多变的特征交叉地编码出诸多形态丰富的具体物种,跨越了漫长时空范畴并构建出复杂多样的演化图景。
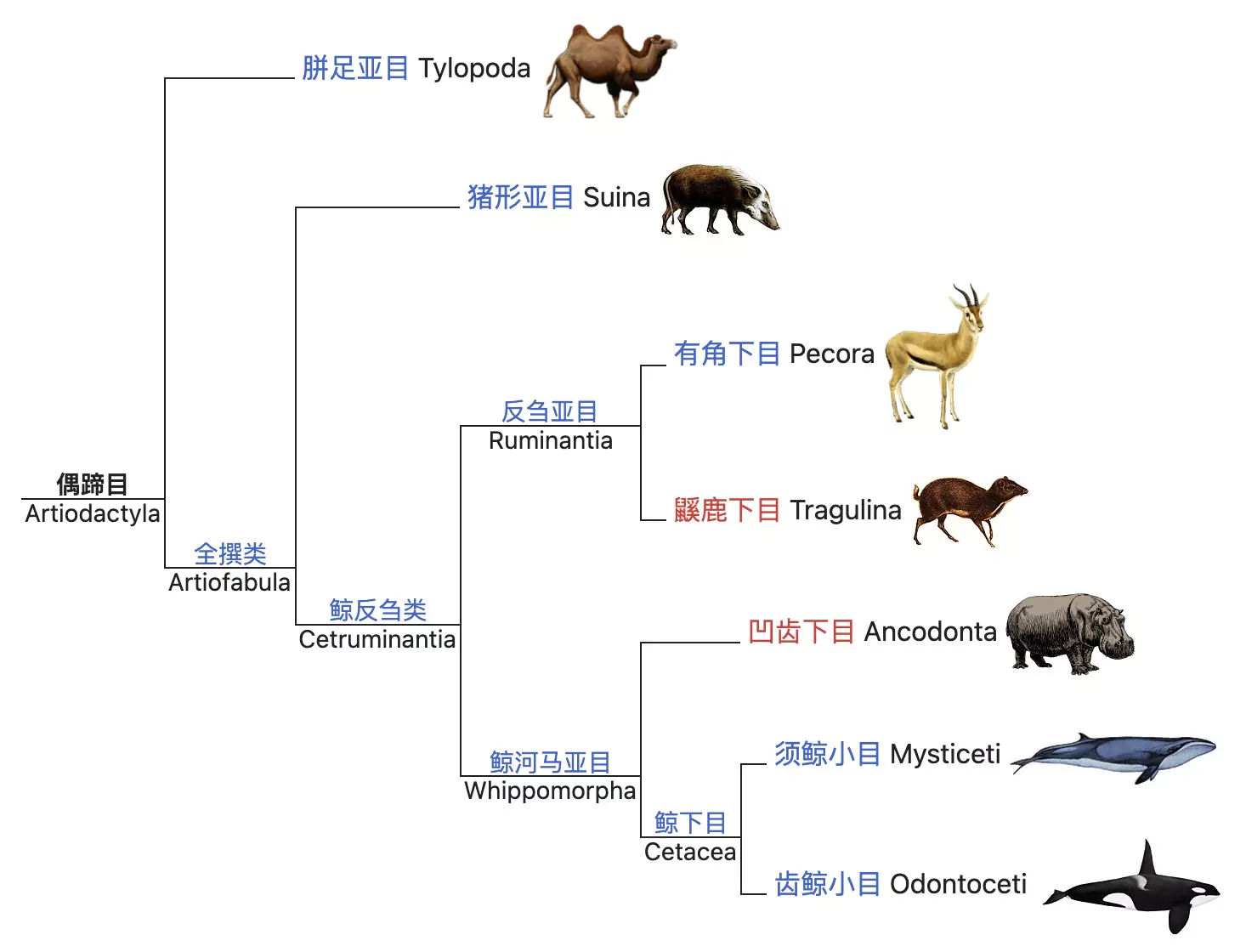
图16. 鲸豚类物种属于传统偶蹄目,但因其四足退化而并不具备偶蹄特征(图源:维基百科)
2.4 门与body plan
在很长一段时间里,界(Kingdom)曾经是生物科学分类法中最高的类别。而随着人们对于细胞分子机制的更深入研究,又在界之上添加了作为最高阶元的域。所有细胞生物被划分为了三个域:古菌域、细菌域、真核域。三域划分也存在一定的争议,可以阅读我们早先的内容【引用葆华的科普:当代生物哲学研讨会(四)探索真核生命的起源之迷:一场演化生物学的壮丽征程】。除此之外,域下面界的划分方式也一直处于争论状态,有“三域五界”或“三域六界”之争。每个界代表着截然不同的基本生命形态,不同界的生物甚至不共享完全一致的密码子元件。到目前为止,动物界和植物界的框架是一直稳定的,只是各种单细胞生物的划分归属问题仍处于前沿研究状态,在此不做过多展开。

图17. 古生物学鼻祖乔治·居维叶(Georges Cuvier,1769年-1832年)(图源:维基百科)
界的下一个分类阶元是门(Phylum)。“门”的这一概念的思想构建最早可以追溯到古生物学鼻祖乔治·居维叶(Georges Cuvier,1769年-1832年)。当居维叶试图对动物界的不同种类生物分门别类时发现,有些动物之间似乎无论如何也找不到具体的共享特征,例如将脊椎动物的任何解剖结构都无法跟昆虫有直接对应关系。于是,当居维叶编写《动物界》一书时,将动物分成了四个分支,即脊椎动物、软体动物、有绞动物(节肢动物和环节动物)和植物型动物(居维叶称作“放射虫”,包含棘皮动物、刺胞动物和其它形式),并且指出这些动物都是独立起源演化出来的,没有任何关联。在这种思想的引领下,海克尔首次提出“门”的概念,将其定义为“一个真正的、完全独立的整体,是其所有下辖物种的集合,这些物种从一个共同的原始形式逐渐进化而来,例如,所有脊椎动物。”
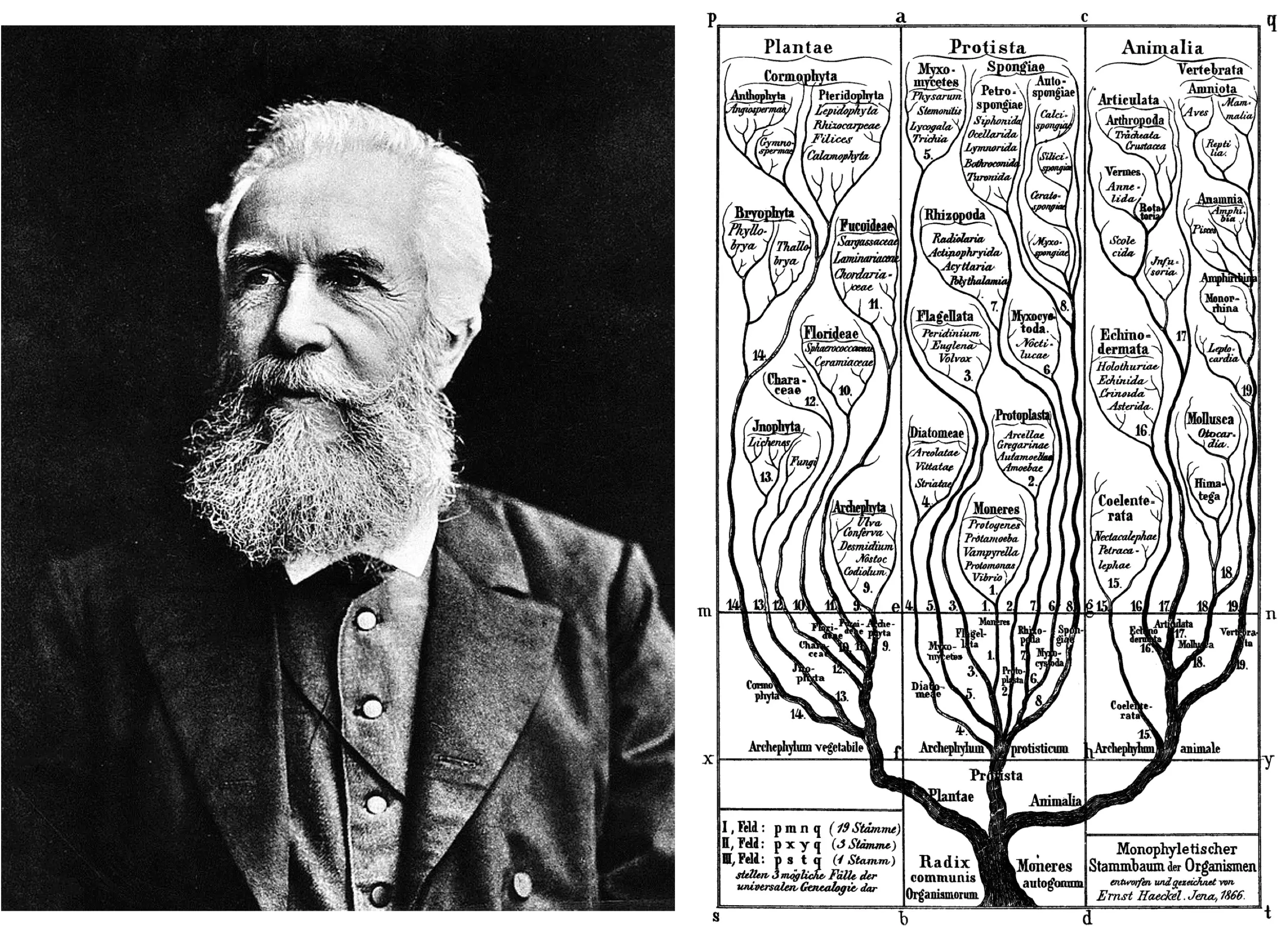
图18. 1866年,德国生物学家恩斯特·海克尔对生命进行三域划分(图源:维基百科)
于是,同一个门的动物共享的基本结构特征组合就被叫做体型呈现(body plan)。定义一个新的门需要强调的是这个门里面的所有生物都无法找到与其他门类生物直接的同源解剖结构。例如,虽然人类和斑马鱼在基本形态上完全不一样,但解剖学会鉴定出一系列同源的骨骼、肌肉、脑区等生理结构,所以只有把它们放在同一个门下面才是合理的。
对于多细胞生物而言,门的划分直接反映的是基础的发育规律。Body plan之所以能在整个门的生物体内都维持保守,是因为它们是产生无数下游特征的结构基础。例如,我们脊索动物门最保守的结构之一就是原肠胚阶段的脊索组织,大多数组织器官的成功分化都需要在脊索的诱导下才能实现,这是发育的因果逻辑决定的。也正因为我们认识到脊椎动物中的脊索结构比脊椎更加基本原始,才将海克尔定义的脊椎动物门变更为脊索动物门,同时将那些无脊椎脊索动物如文昌鱼和海鞘囊括在内。对门类划分的调整都意味着我们对于宏观演化模式有了重大提升,让我们能在更深层次理解演化的原理与内在逻辑。
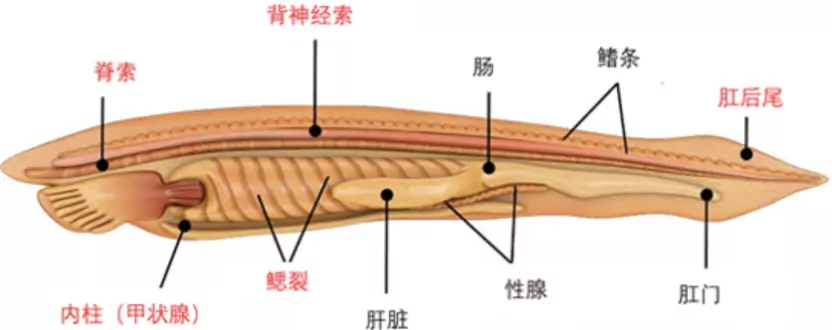
图19. 脊索动物门的body plan(标为红色)(图源:https://biologydictionary.net/vertebrate/)
除了动物界之外,植物界是另一个已被充分研究的领域。几百年来,人们根据维管组织的有无,种子包被方式,花、叶等器官形态划分出了14个现生植物门。
整体上讲,必然是亲缘关系越近的物种大体上长得越像,越能找到更多的类似特征。所以,越是接近科、属级别的划分,越注重具体描述彼此间的形态差异来加以区分;而越接近门、纲级别的划分,则越强调需要用少数保守的共性特征来划分。
也就是说,如果我们发现了一种过去从未见过的生物或化石,哪怕它跟其他所有已知生物相比都看起来长得完全不同,但只要它具备某个门特有的保守特征,我们照样有充分理由把它划分到这个门内。在过去看来,不同门的动植物无法找到直接的同源性证据,但随着发育生物学的深入发展,人们在细胞分化和基因表达调控层面发现不同门生物在解剖结构多样性背后普遍存在深度同源性,这让我们对物种演化有了全新认知。
写在最后
理论上讲,只要得到那棵相对准确的系统发生树(Tree of life),我们完全可以在树的每个二叉节点都命名一个物种类别,这样绝对能保证它们的单系性。当然,真实的物种演化应当并不一定以二叉的形式进行,很有存在多叉或非随机分流等多种复杂状态;所以简单分二叉并进行所有单系群的命名方式必然彻底地丢失了演化的内涵与逻辑。
我们之所以根据脊索等特征定义出脊索动物门,再用脊椎等特征定义出脊椎动物亚门,是因为这样的特征在某个对应的关键祖先节点出现意味着极不寻常的演化事件。在这个过程中,脊椎动物祖先能在某支脊索动物中诞生,是因为脊椎等结构的发育能力需要在脊索组织的诱导下才能构建出来;而脊索动物门属于两侧对称动物则是因为脊索等上游胚胎结构的发育又得建立在两侧对称动物共有的三胚层模式基础上才能实现。因此,我们对于物种门类的划分方式中包含了宏观演化背后的内在逻辑,它不是对于系统发生树进行任意切割都能体现出来的,必须要将树结构和演化趋势进行深度耦合才是有意义的物种分类方式。
总之,界、门、属的概念需要建立在一些生物学普适性原则的基础上提出,而纲、目、科的建立则没有普适性原则。定义一个独立的界需要判断最基本的细胞结构和分子生物学机制的不同,定义一个独立的门需要强调生理形态和解剖结构的不同,定义一个独立的属需要强调对应的生物类群在遗传、演化和生态层面的整体稳定性。
即使存在这些挑战和争议,分类阶元仍然是我们理解和组织生命多样性的重要工具。随着时代的进步,科学技术的不断发展,我们可能会不断审视和调整这些分类阶元的定义与划分。
参考文献
- Löbl, Ivan, et al. “The silent extinction of species and taxonomists—An appeal to science policymakers and legislators.” Diversity 15.10 (2023): 1053.
- Mayden, Richard L. “Consilience and a hierarchy of species concepts: advances toward closure on the species puzzle.” Journal of Nematology 31.2 (1999): 95.
- Richards, Richard A. The species problem: A philosophical analysis. Cambridge University Press, 2010.
- Toews, David PL, et al. “A wood-warbler produced through both interspecific and intergeneric hybridization.” Biology letters 14.11 (2018): 20180557.
- Claridge, Michael F. “Species are real biological entities.” Contemporary Debates in the Philosophy of Biology. Chichester, West Sussex: Wiley-Blackwell (2010): 91-109.
- Mishler, Brent D. “Species are not uniquely real biological entities.” Contemporary debates in philosophy of biology 12 (2010): 110-122.
- Lehman, Hugh. “Are biological species real?.” Philosophy of Science 34.2 (1967): 157-167.
- Morrone, Juan J. “When phylogenetics met biogeography: Willi Hennig, Lars Brundin and the roots of phylogenetic and cladistic biogeography.” Cladistics 39.1 (2023): 58-69.
- Mayr, Ernst. “What is a species, and what is not?.” Philosophy of science 63.2 (1996): 262-277.
- Laurin, Michel. “The PhyloCode: The logical outcome of millennia of evolution of biological nomenclature?.” Zoologica Scripta (2023).
- Cellinese, Nico, David A. Baum, and Brent D. Mishler. “Species and phylogenetic nomenclature.” Systematic Biology 61.5 (2012): 885-891.
- The International Society for Phylogenetic Nomenclature (http://phylonames.org)
- PhyloCode (http://phylonames.org/code).
- Stiller, J., Feng, S., Chowdhury, AA. et al. Complexity of avian evolution revealed by family-level genomes. Nature (2024).

关注 浙江大学生命演化中心公众号 获取中心最新推文